O miosina é um motor molecular, de natureza proteica, capaz de mover-se sobre os filamentos de actina do citosol. A energia que impulsiona o deslocamento da miosina vem da hidrólise do ATP. Por causa disso, a miosina é frequentemente definida como uma enzima mecanoquímica.
Em eucariotos, a miosina é uma proteína muito abundante. Existem diferentes classes de miosina, que são codificadas por uma família de genes. Em leveduras, 5 classes são distinguidas, enquanto em mamíferos dezenas foram descritas..
A miosina tem uma ampla variedade de funções. A miosina I, junto com a actina, participa da movimentação dos ceratócitos.
A miosina II confere rigidez à membrana plasmática, participa da citocinese e da contração muscular. Ambas as miosinas I e II colaboram com a migração celular. As miosinas I e V realizam o transporte das vesículas ao longo dos filamentos de actina.
Índice do artigo
Em micrografias eletrônicas, a estrutura típica das isoformas de miosina tem três domínios: cabeça, pescoço e cauda. Por hidrólise com quimiotripsina, é obtido um segmento constituído pela cabeça e pescoço, denominado meromiosina pesada (HMM), e um segmento da cauda, denominado meromiosina leve (LMM).
O domínio principal é a extremidade N-terminal da cadeia pesada e o domínio cauda é a extremidade C-terminal da cadeia leve.
As classes de miosina podem ser diferenciadas pelo número de cadeias polipeptídicas que as compõem, e pela abundância e classe de cadeia leve anexada ao pescoço.
A miosina I possui uma cadeia polipeptídica, que forma uma cabeça e sua cauda carece de regiões alfa-helicoidais. Considerando que as miosinas I e V têm duas cadeias polipeptídicas e, portanto, formam duas cabeças e uma cauda, na qual as cadeias alfa helicoidais se enrolam para formar uma estrutura semelhante a uma haste.
As miosinas I e V têm locais de ligação para a calmodulina, que regula e liga o Ca+dois, em cadeias leves. Miosina I corrige Ca+dois em cadeias leves, mas é diferente da calmodulina.
No nível mecanoquímico, as miosinas têm três características, a saber:
- A cabeça da miosina é o domínio motor que avança em etapas discretas: a união da cabeça da miosina a um filamento de actina, sua inclinação e posterior separação produzem o movimento da miosina. Este processo é cíclico e depende de ATP.
- Mudanças conformadas: a hidrólise de uma molécula de ATP é acoplada a cada etapa de uma molécula de miosina, por meio de níveis de amplificação e transmissão. Isso envolve grandes mudanças conformacionais da miosina..
O primeiro nível de amplificação é produzido pela perda do grupo gama-fosfato do ATP, que permite uma reorganização dos elementos estruturais no sítio de ligação do ATP. Essa reorganização é coordenada com mudanças estruturais no local de ligação da actina..
O segundo nível de amplificação envolve a comunicação da mudança conformacional no sítio ativo para componentes estruturais do terminal carboxila..
- Direcionalidade: descobriu-se que as miosinas têm polaridade, ou direcionalidade reversa, em direção à extremidade (+) do filamento de actina. Esta conclusão vem dos experimentos de deslizamento de filamento de actina, usando microscópio de luz de fluorescência..
A miosina, junto com a actina, participa da contração muscular, adesão celular, citocinese, enrijecimento das membranas corticais e deslocamento de algumas vesículas, entre outras funções..
Defeitos na miosina podem produzir condições patológicas. Por exemplo, defeitos nas miosinas I e V estão relacionados, respectivamente, a miopatias de miosina e distúrbios de pigmentação (síndrome de Griscelli). Embora distúrbios nas isoformas da miosina VI causem perda auditiva.
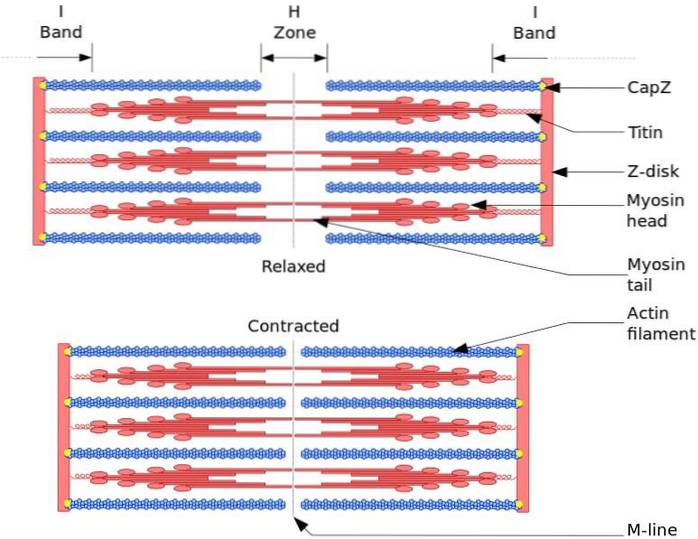
A unidade funcional e estrutural do músculo esquelético é o sarcômero. Durante a contração muscular, o comprimento do sarcômero atinge 30% de seu comprimento original.
Os sarcômeros são compostos de filamentos de miosina grossos e filamentos de actina finos que são organizados de forma complexa. Em geral, as cabeças da miosina estão localizadas nas extremidades distais do filamento e suas caudas em direção ao centro do sarcômero, e a organização é bipolar.
Para que a contração muscular ocorra, as cabeças de miosina nas extremidades opostas devem se mover em direção ao disco Z ou à extremidade (+) do filamento. Como a organização dos filamentos grossos é bipolar, ocorre o deslizamento dos filamentos finos nos filamentos grossos, impulsionado pelo ATP.
A força de deslocamento ocorre porque centenas de cabeças de miosina, de um filamento grosso, interagem com um filamento fino.
Durante a mitose, quando os microtúbulos nos pólos do fuso se separam, a actina e a miosina II formam um anel contrátil no equador da célula. Este anel se contrai, diminuindo seu diâmetro e dividindo a célula em duas partes..
Em células mutantes sem miosina II, a membrana plasmática se deforma facilmente quando uma força externa é aplicada. Isso ocorre porque a miosina II fornece força de agregação às proteínas da membrana plasmática..
No tecido epitelial, os feixes contráteis de actina e miosina II estão localizados nas proximidades da membrana plasmática e formam uma cintura circular que envolve a superfície celular interna. Essa cinta circular determina a forma da célula e mantém o vínculo entre as células..
O contato entre as células ocorre pela união da cinta circular às moléculas de adesão celular, por meio de proteínas de união..
Evidências experimentais revelam que a miosina V realiza o transporte da membrana do aparelho de Golgi para a periferia da célula. Algumas evidências são:
- Nas células do tecido nervoso, por imunofluorescência de astrócitos, verificou-se que a miosina V está localizada próxima ao Golgi..
- Na levedura, as mutações no gene da miosina V interrompem a secreção de proteínas e, consequentemente, a proteína se acumula no citosol.
- As isoformas da miosina I são responsáveis pelo transporte dos vacúolos em direção à membrana celular. Usando anticorpos específicos contra as isoformas da miosina I, verificou-se que essas isoformas estão localizadas em diferentes partes da célula..
Por exemplo, quando uma ameba viva é marcada com um anticorpo contra a miosina IC, o transporte do vacúolo para a membrana é interrompido. Devido a isso, o vacúolo se expande e a célula explode.
Existem vários genes e mutações que causam perda auditiva. Esta doença é frequentemente monogenética .
Mutações não convencionais da miosina, com uma ou duas cabeças de miosina, afetam o funcionamento do ouvido interno. Algumas das isoformas de miosina mutadas são miosina IIIA, miosina VIIA e miosina XVA. Recentemente, duas mutações foram descobertas na miosina VI.
As mutações na miosina VI são c.897G> T e p.926Q. A primeira mutação afeta uma região que interage com o sítio ativo, chamada Switch I. Os homozigotos para a mutação exibem o fenótipo precocemente, causando efeitos graves.
A segunda mutação afeta uma região de resíduos carregados, em uma hélice alfa na cauda da miosina VI. Essa região é importante para a dimerização motora proximal e afeta a função estereociliar da miosina VI..
Outra mutação é p.Asn207Ser, que produz um motor incapaz de produzir força. Isso porque Asn 207 é um resíduo de aminoácido do sítio ativo, cuja função é a ligação e hidrólise do ATP.
A mutação p.Arg657Trp causa perda da função da miosina VI. O resíduo Arg está envolvido nas mudanças conformacionais que acoplam a hidrólise ao movimento da miosina..
Miosina X (Myo10) é uma miosina não convencional que se expressa no cérebro, endotélio e muitos epitélios. Myo10 e três classes de projeções baseadas em actina (filopódios, invadopódios e projeções semelhantes a filopódios) funcionam durante a metástase do câncer.
As células cancerosas invasivas têm um grande número de filopódios e expressam altos níveis de fascina. Esta proteína faz ligações cruzadas entre os filamentos de actina. Para escapar do tumor primário, formam-se invadópodes, ricos em atividade proteolítica, que digerem a matriz extracelular circundante..
Uma vez que as células alcançam a matriz extracelular, as projeções semelhantes a filópodes ajudam a se dispersar e colonizar. Altos níveis de Myo10 indicam alta agressividade e metástase no câncer de mama.
O silenciamento de MyoX causa perda do caráter metastático das células, que são incapazes de formar projeções baseadas em actina. Todas essas projeções têm aderências à base de integrina, que são transportadas pelo Myo10 dentro do filopódio..
MyoX está envolvido na formação do centrossoma. A ausência de MyoX favorece a formação de fusos multipolares. MyoX também está envolvido na sinalização em células cancerosas. Por exemplo, MyoX é ativado por 3,4,5, -inositol trifosfato (PIP3).



Ainda sem comentários