O esfingomielina é o esfingolípido mais abundante nos tecidos animais: a sua presença foi encontrada em todas as membranas celulares estudadas até à data. Apresenta semelhanças estruturais com a fosfatidilcolina em termos de grupo da cabeça polar, razão pela qual também é classificado como um fosfolipídeo (fosfosfingolipídeo).
Na década de 1880, o cientista Johann Thudichum isolou um componente lipídico solúvel em éter do tecido cerebral e chamou-o de esfingomielina. Mais tarde, em 1927, a estrutura deste esfingolipídeo foi relatada como N-acil-esfingosina-1-fosfocolina.
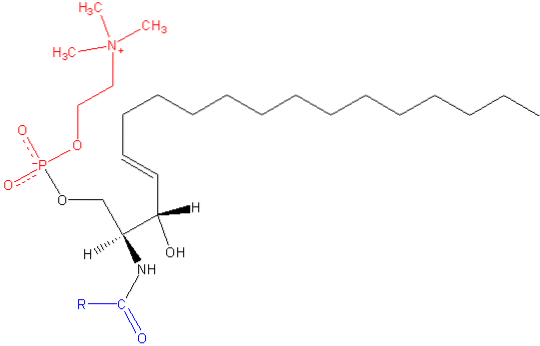
Como os outros esfingolipídeos, a esfingomielina tem funções de sinalização tanto estruturais quanto celulares, e é especialmente abundante nos tecidos nervosos, especificamente na mielina, uma bainha que cobre e isola os axônios de certos neurônios.
Sua distribuição tem sido estudada por meio de experimentos de fracionamento subcelular e degradação enzimática com esfingomielinases, e os resultados indicam que mais da metade da esfingomielina em células eucarióticas é encontrada na membrana plasmática. No entanto, isso depende do tipo de célula. Nos fibroblastos, por exemplo, é responsável por quase 90% dos lipídios totais.
A desregulação dos processos de síntese e metabolismo desse lipídio leva ao desenvolvimento de patologias complexas ou lipidose. Um exemplo disso é a doença de Niemann-Pick hereditária, caracterizada por hepatoesplenomegalia e disfunção neurológica progressiva.
Índice do artigo
A esfingomielina é uma molécula anfipática composta por uma cabeça polar e duas caudas apolares. O grupo da cabeça polar é uma molécula de fosfocolina, por isso pode parecer semelhante à fosfatidilcolina de glicerofosfolipídio (PC). No entanto, existem diferenças substanciais em relação à região interfacial e hidrofóbica entre essas duas moléculas..
A base mais comum em uma molécula de esfingomielina de mamíferos é a ceramida, composta de esfingosina (1,3-dihidroxi-2-amino-4-octadeceno), que tem uma ligação dupla em trans entre os carbonos nas posições 4 e 5 da cadeia de hidrocarbonetos. Seu derivado saturado, esfinganina, também é comum, mas é encontrado em menor extensão.
O comprimento das caudas hidrofóbicas da esfingomielina varia de 16 a 24 átomos de carbono e a composição de ácidos graxos varia dependendo do tecido.
As esfingomielinas da substância branca do cérebro humano, por exemplo, possuem ácido nervônico, as da substância cinzenta contêm principalmente ácido esteárico e a forma prevalente nas plaquetas é o araquidonato..
Geralmente, há uma disparidade de comprimento entre as duas cadeias de ácidos graxos da esfingomielina, o que parece favorecer o fenômeno de "interdigitação" entre os hidrocarbonetos em monocamadas opostas. Isso fornece à membrana estabilidade especial e propriedades particulares em comparação com outras membranas que são mais pobres neste esfingolipídeo..
Na região interfacial da molécula, a esfingomielina possui um grupo amida e uma hidroxila livre no carbono 3, que podem servir como doadores e aceitadores de ligações de hidrogênio para ligações intra e intermoleculares, importantes na definição de domínios laterais e interação. tipos de moléculas.
Os produtos do metabolismo da esfingosina - ceramida, esfingosina, esfingosina 1-fosfato e diacilglicerol - são importantes efetores celulares e lhe conferem papel em múltiplas funções celulares, como apoptose, desenvolvimento e envelhecimento, sinalização celular, entre outras..
Graças à estrutura tridimensional “cilíndrica” da esfingomielina, esse lipídio pode formar domínios de membrana mais compactos e ordenados, o que tem implicações funcionais importantes do ponto de vista proteico, pois pode estabelecer domínios específicos para algumas proteínas integrais de membrana..
As jangadas de lipídios, fases de membrana ou microdomínios ordenados de esfingolipídios, como esfingomielina, alguns glicerofosfolipídios e colesterol, representam plataformas estáveis para a associação de proteínas de membrana com várias funções (receptores, transportadores, etc.).
Caveolae são invaginações da membrana plasmática que recrutam proteínas com âncoras GPI e também são ricas em esfingomielina.
O colesterol, devido à sua rigidez estrutural, afeta significativamente a estrutura das membranas celulares, principalmente nos aspectos relacionados à fluidez, por isso é considerado um elemento essencial.
Como as esfingomielinas possuem doadores e aceitadores de ligações de hidrogênio, acredita-se que sejam capazes de formar interações mais “estáveis” com as moléculas de colesterol. É por isso que se diz que existe uma correlação positiva entre os níveis de colesterol e esfingomielina nas membranas..
A síntese da esfingomielina ocorre no complexo de Golgi, onde a ceramida transportada do retículo endoplasmático (RE) é modificada pela transferência de uma molécula de fosfocolina da fosfatidilcolina, com a liberação concomitante de uma molécula de diacilglicerol. A reação é catalisada por SM sintase (ceramida: fosfatidilcolina fosfocolina transferase).
Há também outra via para a produção de esfingomielina que pode ocorrer pela transferência de uma fosfoetanolamina da fosfatidiletanolamina (PE) para a ceramida, com subsequente metilação da fosfoetanolamina. Acredita-se que isso seja particularmente importante em alguns tecidos nervosos ricos em PE..
A esfingomielina sintase é encontrada no lado luminal da membrana do complexo de Golgi, que é consistente com a localização extra citoplasmática da esfingomielina na maioria das células.
Devido às características do grupo polar da esfingomielina e à aparente ausência de translocases específicas, a orientação topológica desse lipídio depende da enzima sintase..
A degradação da esfingomielina pode ocorrer tanto na membrana plasmática quanto nos lisossomos. A hidrólise lisossomal em ceramida e fosfocolina depende da esfingomielinase ácida, uma glicoproteína lisossomal solúvel cuja atividade tem um pH ótimo em torno de 4,5.
A hidrólise na membrana plasmática é catalisada por uma esfingomielinase que atua a pH 7,4 e que requer íons divalentes de magnésio ou manganês para seu funcionamento. Outras enzimas envolvidas no metabolismo e na reciclagem da esfingomielina são encontradas em diferentes organelas que se conectam entre si por meio de vias de transporte vesicular..



Ainda sem comentários